A)HIV-1 利用宿主因子RVB2调控自身病毒产生
在之前工作中,我们克隆了新宿主抗病毒蛋白RVB2。RVB2特异结合HIV-1 编码结构蛋白Gag的 mRNA上的特定序列,并结合翻译中的Gag蛋白,从而抑制Gag蛋白的翻译,减少病毒颗粒的产生。HIV-1通过自身编码的Env蛋白拮抗RVB2的功能。随后的细胞实验和临床数据表明HIV-1利用这种抑制与拮抗机制减少了不含有囊膜蛋白的病毒颗粒的产生,从而降低了由此带来的机体免疫系统的识别。2015年度,我们深入探究了RVB2抑制Gag翻译的分子机制。研究发现,RVB2通过同时结合翻译过程中的Gag蛋白及其编码mRNA引起mRNA上核糖体停顿。敲低Pelo蛋白减弱RVB2介导的RNA降解,表明RVB2以同no-go decay 相似的机制介导Gag mRNA 的降解。相关成果发表在Cell Host& Microbe,2015。随即,国际同行在Trends Microbiol.杂志的“亮点Spot light”板块配发了对该成果的介绍和评论。
B)M2BP抑制HIV-1病毒颗粒的产生
M2BP是我们实验室筛选到的一个新的宿主抗病毒蛋白。干扰素刺激可上调该蛋白的表达。研究发现在HIV-1的宿主细胞系MT4中,M2BP参与I型干扰素抑制HIV-1病毒复制的过程。M2BP通过抑制HIV-1结构蛋白Gag在细胞膜上的定位从而减少HIV-1病毒颗粒的产生。2015年度,我们深入研究了M2BP作用的分子机制,发现其对病毒的抑制作用依赖于波形蛋白纤维。根据一系列实验结果,我们明确了M2BP抑制HIV-1病毒产生的工作模型:M2BP将HIV-1的Gag蛋白与波形蛋白骨架锚定到一起,从而影响了Gag在细胞膜上的定位,进而抑制了HIV-1病毒颗粒的产生。相关成果正在投稿中。
C)宿主抗HIV-1 因子iVIP的鉴定和机制分析
通过对部分干扰素上调基因进行筛选,我们得到宿主抗HIV-1因子iVIP。 iVIP是一个膜蛋白,膜定位对其抗病毒活性至关重要。iVIP表达不影响细胞内病毒蛋白的表达,但显著降低上清中病毒颗粒的含量,表明iVIP对HIV-1的抑制发生在装配或出芽释放阶段。进一步研究发现,iVIP的抗病毒活性可以被病毒编码的包膜蛋白Env拮抗。对iVIP的深入研究将进一步揭示病毒与宿主间复杂的相互作用。
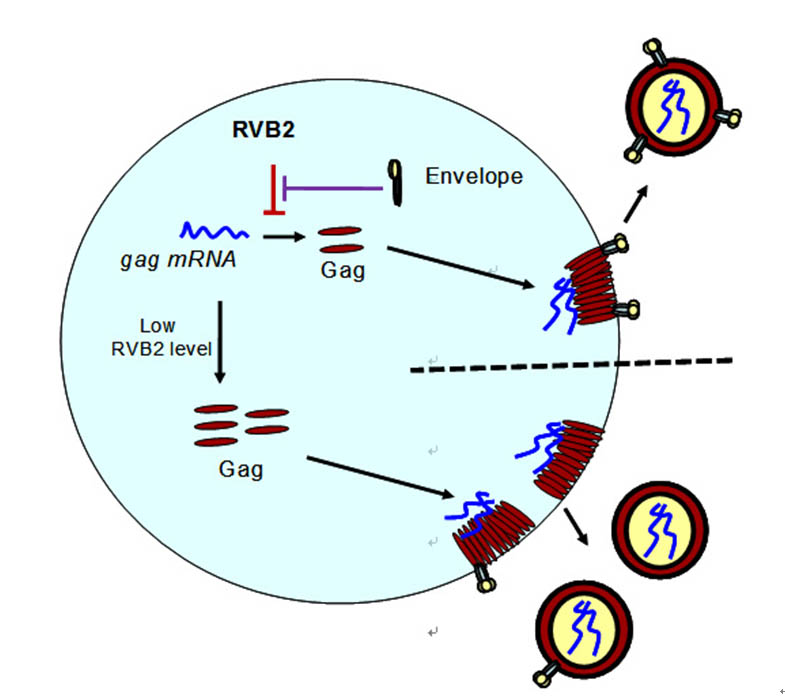
图示:RVB2与HIV-1相互作用的模式图。在病毒产生早期,Envelope(Env)蛋白的表达量很低,内源的RVB2有效控制了病毒Gag的表达,从而降低了病毒颗粒的产生。随着HIV-1 Env蛋白的积累,产生了有效拮抗RVB2的条件,Gag的表达不再被抑制。通过该机制,保障了Gag 蛋白与Env蛋白的比例和可感染性病毒颗粒的产生;敲低内源RVB2, Gag蛋白大量表达,而没有足够多的Env蛋白匹配,产生大量不含或仅含有少量Env的病毒, 减弱了病毒粒子的感染力。

