 2021年科研进展
2021年科研进展2021年11月24日,金沙集团1862cc许瑞明研究团队在《Genes & Development》杂志在线发表了题为"Distinct histone H3-H4 binding modes of sNASP reveal the basis for cooperation and competition of histone chaperones"的研究论文,揭示了组蛋白伴侣sNASP-ASF1结合组蛋白H3-H4的协同与竞争机制,为理解组蛋白H3-H4的结合模式及其在组蛋白伴侣网络间的传递提供了新的视角。
组蛋白伴侣作为调控染色质组装的重要因子,与真核细胞表观遗传信息的提取与传递紧密相关,其功能异常会影响染色质正确组装,进而影响DNA复制与基因转录、基因组稳定和细胞重编程等生物学过程。团队先前的研究解析了组蛋白伴侣HJURP特异性识别着丝粒区组蛋白变体CENP-A的结构机理(Genes & Dev, 2011);揭示了组蛋白变体H3.3被其伴侣蛋白DAXX精确识别的结构基础(NSMB, 2012),以及阐明了Asf1调控组蛋白乙酰化修饰的分子机制(Cell, 2018)。但对于另一类伴侣蛋白sNASP如何在细胞中结合和存储组蛋白H3-H4以及组蛋白H3-H4如何在伴侣网络中接力传递,其分子机制尚不清楚。此次成果是在该研究方向的一个重要进展。
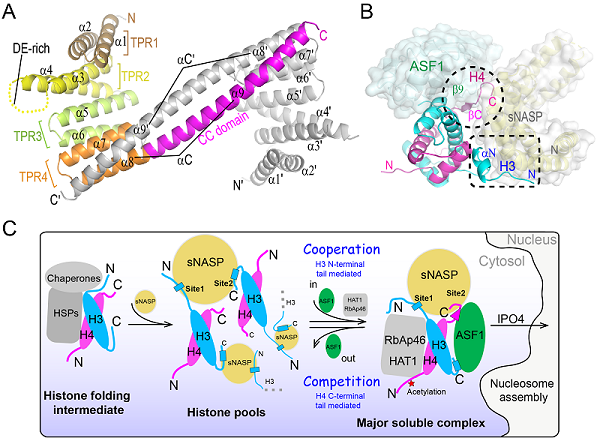
图1 人源组蛋白伴侣sNASP-ASF1协同与竞争结合组蛋白H3-H4的结构基础
(A)sNASP TPR结构域二聚体的晶体结构;(B)sNASP-ASF1-H3-H4四元复合物的晶体结构;(C)组蛋白H3-H4在伴侣网络中接力传递的模式图
该研究首先发现人源组蛋白伴侣sNASP在体外同时存在单体和二聚体两种形式,并解析了sNASP TPR结构域二聚体2.9Å的晶体结构。该结构显示sNASP的TPR4 motif中的α8和Coiled-Coil domain(α9)形成一个超长螺旋αC,介导了sNASP二聚体的形成(图1A)。虽然αC不参与结合组蛋白H3-H4,但缺少该段结构域,将严重影响核小体的组装。接着团队解析了sNASP-ASF1-H3-H4四元复合物3.0Å的晶体结构,复合物结构显示sNASP以单体形式存在于复合物中,介导sNASP二聚体形成的超长螺旋αC不再与邻近分子配对,而是一分为二(α8和α9),并以180度大转弯折回与自身配对;ASF1通过底物组蛋白H3-H4作为媒介,与sNASP间接作用;sNASP与组蛋白H3-H4之间的相互作用位点主要有两处:H3的N端尾巴中的αN和H4的C端尾巴(图1B)。结合体外生化实验和体内细胞荧光共定位技术,团队发现在ASF1存在的情况下,H3的N端尾巴对于sNASP的结合至关重要。H4的C端尾巴虽然对于sNASP的相互作用只起到辅助作用,但有意思的是,当缺少H4的C端尾巴时,ASF1将被踢出去,形成sNASP-H3-H4三元复合物。这提示H4的C端尾巴可以介导组蛋白H3-H4在sNASP-ASF1之间的传递。此外,研究还发现在没有ASF1的情况下,sNASP以一种全新的方式结合组蛋白H3-H4。此时的组蛋白H3-H4主要通过其核心区域与sNASP的TPR4 motif以及位于TPR2 motif中间的酸性loop区域存在相互作用。最后团队根据该论文实验结果并结合先前的研究,提出了组蛋白H3-H4在伴侣网络中接力传递的模式图(图1C),该图全面展示了细胞质中新合成的组蛋白H3-H4通过一系列伴侣蛋白的接力传递,最终进入细胞核进行染色质组装的途径。
金沙集团1862cc许瑞明研究员和刘超培副研究员是本研究论文的共同通讯作者。许瑞明课题组刘超培副研究员和金文星副研究员为本文的共同第一作者。金沙集团1862cc李国红研究员以及其课题组博士生胡杰,安徽大学王明珠教授和许瑞明课题组陈晶晶工程师对本论文都有重要贡献。该研究获得了国家自然科学基金委、科技部重点研发计划、中科院战略性先导科技专项和金沙集团1862cc青年创新促进会的资助。
文章链接:http://www.genesdev.org/cgi/doi/10.1101/gad.349100.121
(供稿:许瑞明研究组)
 附件下载:
附件下载: