 2022年图片新闻
2022年图片新闻自噬是指通过形成双层膜结构的自噬体,包裹部分胞质并运送到溶酶体进行降解及回收的过程,对抵抗各种应激和维持细胞稳态至关重要。自噬体形成的关键步骤包括隔离膜(自噬体前体)的启始、成核、延伸以及闭合。人们对自噬体形成分子机制的了解主要来自对单细胞酵母自噬的研究。多细胞生物自噬体的形成过程更加复杂,包括多个独特的步骤以及多细胞生物特有的自噬蛋白参与。多细胞生物自噬与酵母自噬的一个重要区别在于自噬体形成的位置。酵母自噬体在液泡膜上形成,而多细胞生物自噬体在内质网的多个位点同时形成。鉴定决定自噬体在内质网上形成的信号是自噬领域一个长期悬而未决的科学问题。
2022年10月5日,金沙集团1862cc张宏课题组在《Cell》杂志在线发表了题为"Calcium transients on the ER surface trigger liquid-liquid phase separation of FIP200 to specify autophagosome initiation sites"的研究论文。该文发现:自噬诱导条件下引起的内质网表面的钙瞬变是决定自噬体在内质网上形成的关键信号。内质网表面的钙瞬变引起参与自噬起始的FIP200复合物发生液-液相分离,形成的FIP200凝聚体进而与内质网膜蛋白VAPs和ATLs结合定位于内质网,成为自噬体起始位点。
研究者首先发现钙离子快速螯合剂BAPTA-AM可以抑制参与自噬起始的FIP200复合物在内质网上形成凝聚体,但这一过程不能被慢速钙离子螯合剂EGTA-AM阻断。这提示快速的局部的钙离子变化,而非稳态的钙离子浓度变化,可能参与了自噬起始过程。研究人员构建了内质网跨膜结构域CYB5与快速钙离子探针GCaMP6f的融合蛋白,来检测自噬诱导下内质网外膜表面钙离子浓度的变化。多模态超分辨活细胞成像(Multi-SIM)显示,饥饿或Torin1处理等自噬诱导条件下,内质网外膜发生钙瞬变,这些钙信号能被BAPTA-AM阻断却不能被EGTA-AM阻断。
进一步研究表明,内质网表面的钙瞬变触发参与自噬起始的FIP200复合物发生液-液相分离,形成的FIP200凝聚体通过与内质网膜蛋白VAPs和ATLs结合,定位于内质网上,该结构即为自噬体起始位点。研究还发现,张宏课题组前期鉴定的内质网定位的新自噬蛋白EPG-4/EI24调控内质网表面钙瞬变的幅度、频率和持续时间。在EI24敲除细胞中,内质网表面呈现出持续的钙瞬变,导致FIP200复合物在内质网上异常累积。通过化学试剂处理,或敲低内质网表面钙通道的活性,可以降低EI24敲除引起的钙瞬变并拯救其自噬缺陷的表型。
综上,该研究发现在自噬诱导条件下,内质网表面发生的钙瞬变诱导自噬起始FIP200复合物发生液-液相分离。然后FIP200凝聚体与内质网膜蛋白VAPs和ATLs结合并稳定定位于内质网上,成为自噬起始位点。该工作揭示了内质网表面钙瞬变是决定自噬体在内质网上形成的关键信号,极大地促进了我们对多细胞生物自噬分子机制的理解。
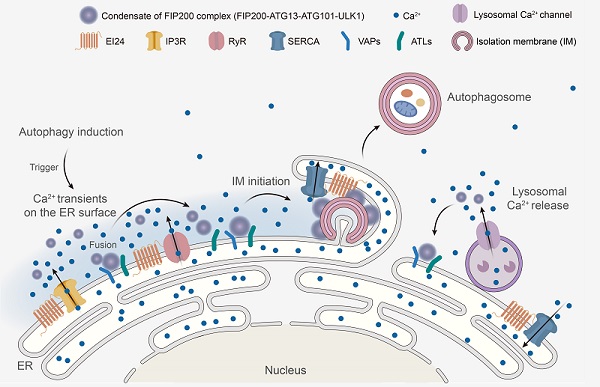
图. 内质网外膜钙瞬变触发FIP200复合物发生液-液相分离,并形成内质网上自噬起始位点的模式图
中科院生物物理所张宏研究员为本文的通讯作者。张宏课题组助理研究员郑巧霞和助理研究员陈永为本文共同第一作者。该研究获得国家自然科学基金、国家重点研发计划资助。生物物理所成像平台为该研究提供了重要的技术支持。
文章链接:https://www.cell.com/cell/fulltext/S0092-8674(22)01123-0#%20
(供稿:张宏研究组)
 附件下载:
附件下载: